
A new study published in Evolution Letters investigates why metabolic rates are so variable between organisms. Here, lead author Dr Amanda Pettersen tell us what she found.
Metabolic rate – the rate at which organisms take up, transform, and expend energy, is linked with the “pace-of-life”. Individuals with high metabolic rates often grow faster, reproduce sooner, and are shorter lived, compared with individuals with low metabolism that are often longer lived. Differences in energy expenditure among individuals are likely to result in differences in survival and reproduction, so we expect metabolic rates are under selection. Evolutionary theory predicts that over time, selection should deplete variation in traits, yet even after accounting for body size and temperature, we can observe a threefold difference in metabolic rates across individuals of the same species. How can we account for this variation?
The presence and magnitude of biotic and abiotic factors can vary considerably in natural environments, even across small spatial scales. Differences in selection regimes across environments may maintain variation in metabolic rates, even under strong selection in any single environment. This is because a fast or slow pace-of-life might be advantageous depending on the environment an organism inhabits. While variation in selection over large and small spatial scales have been found previously, field estimates of selection on metabolism across environments are historically rare.

To investigate the role of environmental variation in maintaining trait variation, we studied the model marine invertebrate, Bugula neritina. The adult stage of Bugula neritina is a colonial bryozoan, comprised of thousands of individual subunits (zooids). Some zooids contain reproductive structures (ovicells) that hold individual larvae. In response to light exposure at dawn, the parent colony releases hundreds of swimming larvae into the plankton – the larvae soon find a suitable substrate to settle on, such as a pier pylon, and metamorphose into the first zooid of the colony. From here, the zooids begin budding and the colony grows, forming ovicells approximately four weeks after settlement.
We settled individual marine bryozoan larvae by pipetting them in a drop of water onto small pieces of acetate (as a settlement substrate) and measured their metabolic rates. We measured metabolic rate (rate of oxygen consumption) at two time points: 2 hours (“Metabolic Rate Early”; MRE) and 24 hours (“Metabolic Rate Late”; MRL) post-settlement. We then glued the settlers on the acetate pieces onto tiles in one of three competition environments: no competition (“nocomp”), within-species competitors (“intra”), and a pre-established marine community (“inter”). We randomly assorted these treatment tiles onto 10 backing panels (36 tiles per panel) and hung them at 1.5m depth in the subtidal at our field site – a marina in Melbourne, Australia. We monitored individuals approximately weekly for 4 months, measuring fitness: survival (“viability”), the ability to reproduce (“fertility”), and reproductive output (“fecundity”), along with our pace-of-life traits: growth, age at onset of reproduction, and longevity.
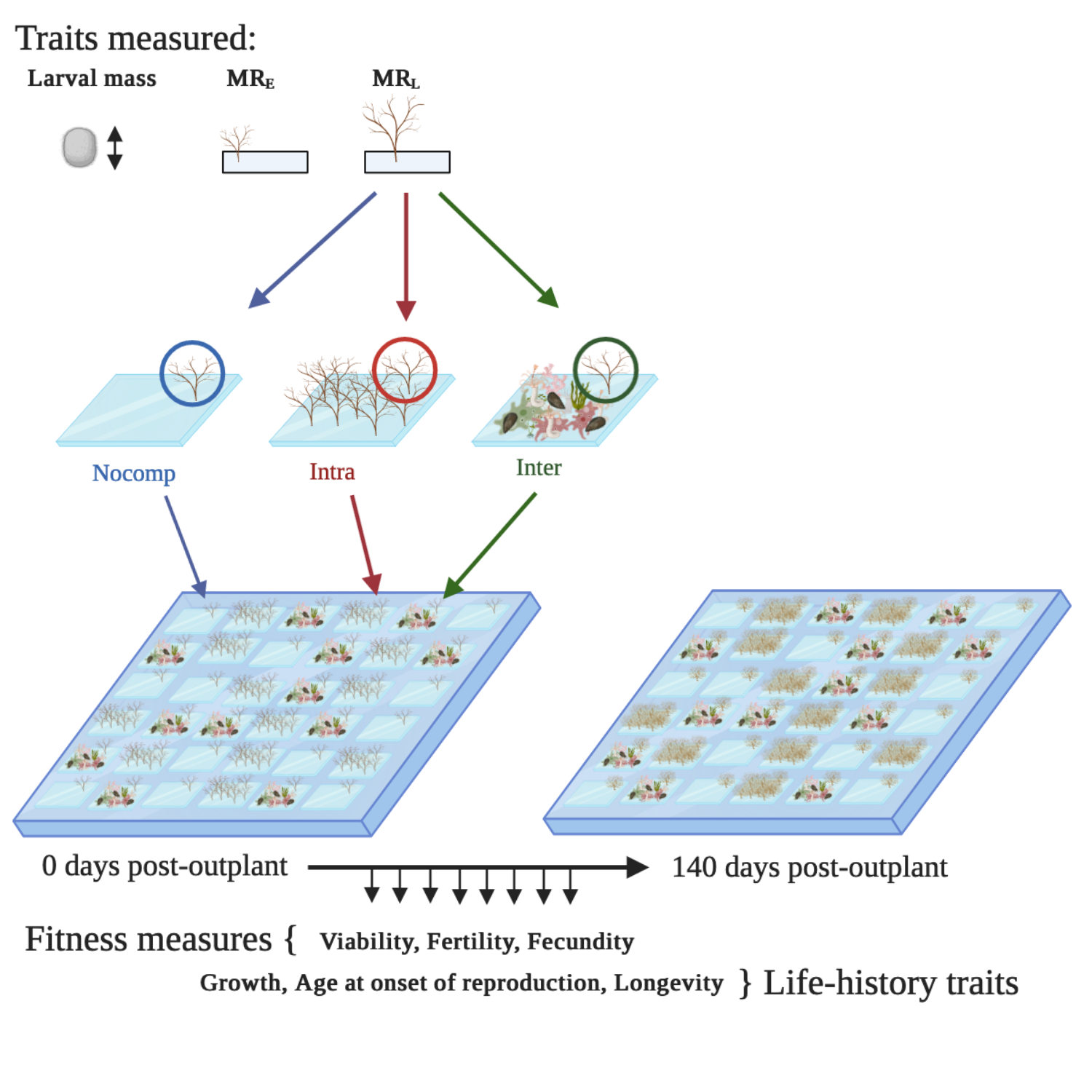
We found that selection on metabolic rates (MRE and MRL) varies among competition environments separated by only a few centimetres – competition selects for a faster pace-of-life, compared with competition-free environments. High-metabolism individuals are better able to withstand intense competition from both within-species competitors and in a pre-established marine community. However, low-metabolism individuals live longer, and showed higher fitness under competition-free conditions. If arriving into a competition-free space, individuals will have higher fitness if they grow slow and reproduce later but live longer. Hence, the environment-dependent nature of selection on metabolism and the pace-of-life is likely to maintain variation in metabolic rates.
These findings support previous groundwork in field ecology, suggesting that differences in selection regimes maintain trait variation. We found that selection on metabolic rates can vary considerably, even over a few centimetres! A high metabolic rate and fast pace-of-life is favoured under strong competition – individuals die young but reproduce sooner. However, in the rare case that individuals find themselves in a competition-free environment, then a low metabolic rate and slow pace-of-life can provide a fitness advantage.
Dr Amanda Pettersen is a postdoctoral researcher at the University of Lund. The original article is freely available to read and download from Evolution Letters.
You can also watch a short talk by Dr Pettersen on the study here:

