
The findings of a new study published in Evolution Letters help explain the coexistence of many closely related flowering plant species, in one of the most diverse temperate ecosystems in the world. Lead author Dr Alex Skeels tells us more.
Flowers are some of the most beautiful and fascinating structures in nature and most people can recognise a handful of plant species just from the sight of their flowers (think roses, or Banksias if you’re Australian). In fact, most plant species’ flowers are so unique that they are among the most important features used by taxonomists and naturalists to identify species in the field. Angiosperms, or flowering plants, are the most diverse group of plants on the planet, and it has long been suspected that the huge diversity in flower size, shape, colour, and scent, may explain this. So, understanding how flowers evolve over time is key to understanding the origin of plant biodiversity.
Natural selection is the great evolutionary force that shapes complex structures like flowers, but several different agents of selection may be involved in shaping the diversity of flowers. The main role of flowers is to attract pollinators to share pollen with individuals of the same species. But selection might push flowers of different populations of a species to diverge when, for example, populations benefit from attracting different pollinators in different ecological communities. This can lead to speciation as the diverging populations no longer share pollen and therefore no longer reproduce sexually.

In other cases, individuals of different species might have similar flowers, and would be expected to attract similar pollinators. This might not matter too much if there are plenty of pollinators available, if there is not a strong cost to sharing pollen, or those species do not occur together in the same communities, and flowers might remain the same or change in response to changes in local pollinators over time. But as soon as these conditions change and competition for pollinators between species with similar flowers increases then we expect natural selection to favour divergence.
These examples differ in whether flower divergence drives speciation, or whether flower divergence comes later, either drifting in response to local changes in pollinator communities or driven by competition and other negative interactions between species. This is an important distinction in understanding whether flower divergence is a cause or a consequence of plant biodiversity. But how do we tell these scenarios apart? In this study we came up with some predictions of what we would expect under these three different scenarios. We tested these predictions in Hakea species from Southwest Australia.
Hakea are a fascinating genus of plants endemic to Australia. They show exceptional diversity in the Mediterranean-climate region of Southwest Australia which is a recognised biodiversity hotspot due to its high diversity of endemic plants under threat from human land-use modifications, particularly from agriculture (the region’s known as the wheat-belt). The region is famous for its fairly extreme environment (poor soils and hot dry summers) and super-rich plant communities, and sitting a bit below a similar Mediterranean-type ecosystem, the Cape of South Africa, is one of the most diverse temperate ecosystems in the world. Hakea are generally known as an unfriendly genus of spiky shrubs and perhaps less well recognised for their diversity of beautiful flowers, especially compared to their iconic relatives in the Proteaceae family – the Banksias. Previous studies of Hakea by Lamont and colleagues have shown flower size and colour to be important traits in determining pollinator specificity in the genus, so we went about collating data and characterising these traits to test our model predictions.

To do this, we decided to make use of as much readily available data as possible and this meant using digital photographs to characterise flower colour. We used images from the Australian Plant Image Index (APII, anbg.gov.au/photo), a resource provided by the Australian National Botanical Gardens in Canberra (just next to the Australian National University where the authors were based), as well as from iNaturalist (inaturalist.org) and even Flickr (flickr.com) when no other sources had photographs of particular species. Our photographic library therefore had a collection of photos from professional botanists alongside amateur naturalists and citizen scientists, and represents a non-traditional approach to data collection that is increasingly becoming a powerful resource to support big-data analyses where field-based data collection is not always feasible. From the photos we could extract values for different organs in the flower (for example pistils and perianths, see below) to test our model predictions.
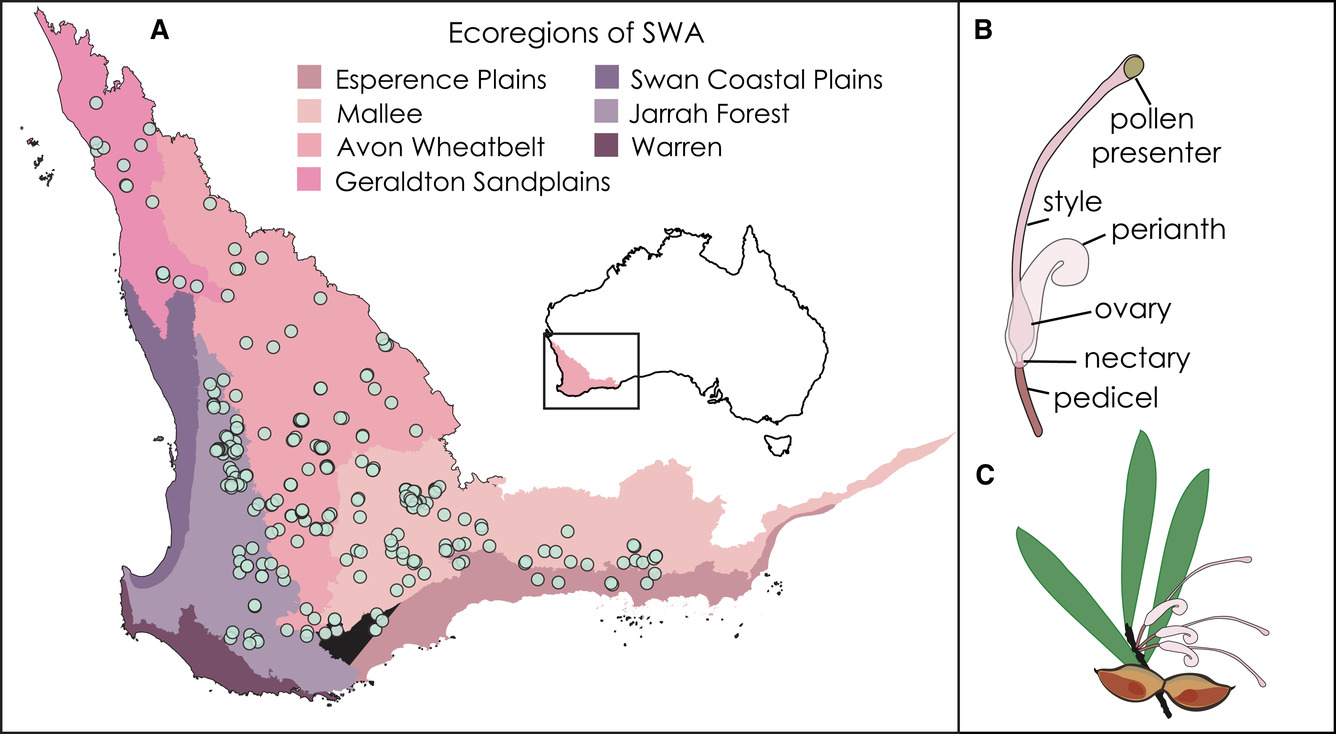
We found that Hakea species typically co-occur with other Hakea species with very different flower colours, which suggests natural selection is preventing species with similar flowers from co-existing. We also found that, despite the genus being over 35 million years old, flower colour evolved rapidly from the Mid-Miocene about 15 million years ago and has sped-up towards the present-day. These patterns fit our predictions expected from a competition model of flower evolution rather than a speciation model. This makes sense because in Southwest Australia many closely-related species are packed into communities and we expect competition to be rampant and a strong selective force. Yet this result also in some ways goes against the traditional view that flower divergence is most important during speciation and while this might be true in many cases, we found that, at least for flower colour, divergence seems to come subsequent to speciation, as biodiversity and the number of potential species interactions accumulated in Southwest Australia. Altogether, the interesting history of flower evolution in Hakea helps shine a light on the processes driving the incredible diversity of flowers and flowering plants.
Dr Alex Skeels is currently a post-doc at ETH Zürich and the Swiss Federal Research Institute for Forest, Snow and Landscape (WSL). The above work was carried out as part of Alex’s PhD with the Macroevolution and Macroecology Group from the Australian National University and Behavioural Ecology and Macroevolution Group from the University of Melbourne. The original study is freely available to read and download from Evolution Letters.

